- Hernandez Hernandez Myriam Yamilli
- Hernandez Ocejo Ingrid Yazmin
- Meza Castillo Cindy Yohali
- Quiroz Cobos Sharon Lizbeth
- Garcia Cervantes Clara Lizeth
- Estevez Bautista Rosa Irene
 |
EL CITOESQUELETO Y LA MOTILIDAD CELULAR MICROTUBULOS, MICROFILAMENTOS, FILAMENTOS INTERMEDIARIOS.
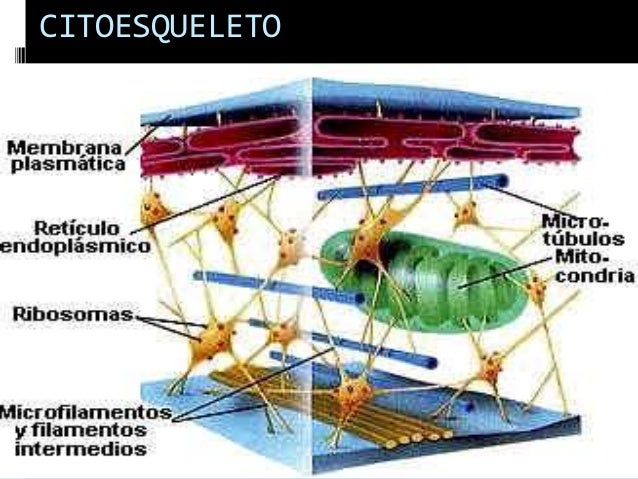
El citoesqueleto es propio de las células
eucarióticas. Es una estructura tridimensional dinámica que se extiende a
través del citoplasma. Por lo tanto la idea de que el citoplasma de la célula
es una masa amorfa y gelatinosa es equivocada.
Esta matriz fibrosa de proteínas se extiende por
el citoplasma entre el núcleo y la cara interna de la membrana plasmática,
ayudando a definir la forma de la célula e interviniendo en la locomoción y
división celular. Es decir que el citoesqueleto no sólo da estabilidad a la
célula como un esqueleto, sino que es también como el músculo interviene en el
movimiento celular. Por lo tanto podríamos llamarlo también “citomusculatura”.
Podemos agregar que el citoesqueleto condiciona el movimiento de las organelas
del interior de la célula y tiene gran importancia metabólica, dando un
andamiaje a los procesos moleculares que se realizan en el citoplasma.
El citoesqueleto es característico de las
células eucariontes ya que ESTA AUSENTE EN LOS PROCARIONTES. Por lo que podría
ser un factor esencial en la evolución de los eucariotas
De esta forma podemos enunciar las siguientes funciones del citoesqueleto:
Ø Estabilidad celular y forma celular
Ø Locomoción celular
Ø División celular
Ø Movimiento de los orgánulos internos
Ø Regulación metabólica
Ø Locomoción celular
Ø División celular
Ø Movimiento de los orgánulos internos
Ø Regulación metabólica
1. CITOSOL, ERGASTOPLASMA Y CITOESQUELETO
 Al realizar el
fraccionamiento celular, una vez separadas las fracciones nucleares, mitocondrial y micro soma, se
logra la fracción sobrenadante, soluble o citosol que es la que contiene
las proteínas y enzimas solubles de la matiz citoplasmática. Estas constituyen
del 20-25% de las proteínas totales de la célula. Entre las enzimas solubles
importantes, presentes en la matriz, se hallan las que interviene en la
glucólisis y en la activación de los aminoácidos para la síntesis de proteínas.
En esta fracción se encuentra también las enzimas para múltiples reacciones que
requieren ATP y, además, el ARN de transferencia o soluble y toda la maquinaria
para la síntesis de proteínas, con inclusión de los ribosomas.
Al realizar el
fraccionamiento celular, una vez separadas las fracciones nucleares, mitocondrial y micro soma, se
logra la fracción sobrenadante, soluble o citosol que es la que contiene
las proteínas y enzimas solubles de la matiz citoplasmática. Estas constituyen
del 20-25% de las proteínas totales de la célula. Entre las enzimas solubles
importantes, presentes en la matriz, se hallan las que interviene en la
glucólisis y en la activación de los aminoácidos para la síntesis de proteínas.
En esta fracción se encuentra también las enzimas para múltiples reacciones que
requieren ATP y, además, el ARN de transferencia o soluble y toda la maquinaria
para la síntesis de proteínas, con inclusión de los ribosomas.
Mientras que al principio se expreso que el citosol era
esencialmente amorfo, a fines del siglo XIX se descubrió que algunas porciones
de citoplasma de ciertas células tenian
propiedades especiales de coloración. Como estas zonas se coloreaban con los
colorantes básicos al igual que el núcleo, recibieron el nombre de citoplasma basofilo o cromidial (Hertwig).
La denominación ergastoplasma (del grupo ergazomal, elaborar y transformar),
que aún se usa en la actualidad, fue acuñada por Gammier en 1887, para signifar
que los biosíntesis es la función fundamental de esta sustancia.
El
ergastoplasma incluye las regiones basòfilas del citoplasma, como los cuerpos
de NIssl de las neuronas, el citoplasma basal de las células serosas (por
ejemplo, células secretoras del páncreas
y de la parótida, y células principales del estomago) y los grupos
basòfilos del hígado. Caspersson, Brachet
y otros demostraron que la intensa basofilia del citoplasma se debe a la
presencia del ácido ribonucleico. En efecto, el citoplasma pierde sus propiedades tintóreas cuando la
célula es tratada con ribonucleasa, una enzima que hidroliza el ARN.
El ARN se encuentra principalmente en los ribosomas y, en consecuencia, se
dedujo una relación entre el ergastoplasma
y la síntesis de proteínas.
El nombre citoesqueloto
fue acuñado hace muchos años, pero
luego se abandonó porque muchas de las estructuras citoplasmáticas se
consideraron como artificios de fijación. En 1928 Koltzoff consideró la existencia de
una organización fibrosa en la estructura del protoplasma. Afirmó “ que cada
célula es un sistema de componentes líquidos y de esqueletos rígidos que generan la forma (de la célula), y aunque
rara vez se observan fibras esqueléticas en las células vivientes o fijadas,
esto solo significa que estas fibrillas son muy finas y no se distinguen por su
índice de refracción de la solución coloidal que las rodea”. Por lo tanto,
concibió un citoesqueleto que determina la forma celular y sus cambios. Confirmando esta suposición
el microscopio electrónico ha relevado
que la mayoría de las células eucariotas ha revelado que la mayoría de las
células eucariotas tiene una trama cito esquelética formada por micro túbulos,
micro filamentos y filamentos intermediarios.
Se han hecho grandes progresos en el aislamiento de las
proteínas que constituyen esta rama cito esquelética ( tubulina, actina,
miosina, tropomiosina y otras células relacionadas) , asi conocimiento del
mecanismo de organización de èstas. Por otra parte, mediante el uso
de anticuerpos específicos se ha podido seguir bajo el microscopio la
disposición de los microtùbulos y
microfilamentos. La microscopia electrónica de alto montaje ha demostrado a su
vez que hay una trama tridimensional en la matriz citoplasmatica. En la actualidad se cree que las diversas formas de motilidad celular
son el resultado de la interacción entre las diversas clases de microfilamentos
y los microtùbulos que se encuentran en la matriz citoplasmantica. El sistemas
más estructurado està represando por la
miofibrilla del mùsculo esquelético, en el que hay mecanismo
macfromolecular muy adaptado a la contractilidad. Sin embargo es ahora evidente
que las células no musculares usan mecanismos semejantes.Aquí describimos
los microtubulos y microfilametos, poniendo énfasis sobre el papel que
desempeñan en el citoesqueleto y en las diversas formas de motilidad celular.
Resuemen
El Citoesqueleto
La matriz citoplasmática o citosol representa el medio interno de la
célula. En el citosol tiene lugar muchas funciones relacionadas con las
transformaciones sol-gel (ciclosis,
movimiento ameboide, formación del huso, clivaje celular). En él se encuentran
las enzimas glucoliticas y toda la maquinaria para la síntesis proteica. Las
regiones basòfolicas del citosol son ricas en ARN (ribosomas), y anteriormente
fueron denominadas ergastoplasma. La denominación de citoesqueleto
se aplicó a la trama de microtùbulos, mcrofilamentos y filamentos
intermediarios que atraviesa el citosol y que está relacionada con las varias
formas de motilidad celular. Las principales proteínas presentes en el
citoesqueleto, es decir: tubulina (microtubulos) y actina, miosina,
tropomiosina y otras (microfilamentos), también forma parte del músculo. Por lo tanto, las mimas proteínas intervienen en
la contracción del músculo y de las células no musculares. 2. MICROTUBULOS
 Los microtubulos son estructuras universalmente
presentes en el citoplasma de los eucariontes, que se caracterizan por su
aspecto tubular y por las constancias de sus propiedades en los diferentes
tipos celulares.
Los microtubulos son estructuras universalmente
presentes en el citoplasma de los eucariontes, que se caracterizan por su
aspecto tubular y por las constancias de sus propiedades en los diferentes
tipos celulares.
La mayoría de los microtubulos son muy lábiles y no
resisten los efectos de agentes filtradores como el tetroxido de osmio. Por
ellos, solo a partir de 1963, con la introducción del glutaradehido como
fijador para microscopia electrónica, pudo intensificarse el estudio de estas
estructuras.
Las primeras observaciones de estructuras tubulares
fueron realizadas por de robertis y franchi, en 1953, el el axoplasma separando
fibras nerviosas mielinicas, donde los neurotubulos aparecen como elementos
cilíndricos alargados, no ramificados, de 20 a 30 nm de diámetro y de longitud
indefinida los microtubulos se observaron posteriormente en gran variedad
células animales
Los microtubulos citoplasmáticos son de tamaño
uniforme y notablemente rectilíneos. Tienen alrededor de 25nm de diámetro
externo y varios micrones de longitud. En los cortes transversales presentan
una configuración anular, con una pared densa de unos 6nm de espesor, y un
centro más claro. Cada microtubulo está rodeado por una zona de baja densidad
electrónica, que carece de ribosomas, lineales o espirales, de un diámetro
aproximado de 5nm, compuesto a su vez por subunidades. En un corte trasversal
se encuentra 13 subunidades, separadas por 4,5nm entre el centro y centro. El
empleo de técnicas de coloración negativa ha permitido demostrar que los
microtubulos tienen luz y que pared está constituida por subunidades.
Ocasionalmente se han detectado bastones de algunos microtubulos
Aunque todos los microtubulos estudiados tienen
aproximadamente las mismas características morfológicas, es evidente que
difieres de sus propiedades. Por ejemplo, los microtubulos de ciclios y
flagelos son muchos más resistentes a diversos tratamientos; en cambio, los que
constituyen las fibras del huso y los que se encuentran en el citoplasma sueles
ser estructuras lábiles y transitorias, los micritubulos citoplasmáticos
desaparecen a 0º C y al ser tratados con colchicima.
LA TUBULINA,
PROTEINA DE LOS MICROTUBULOS
Los
microtubulos están compuestos por subunidades proteicas, que son bastantes similares
en gran variedad de tipos celulares. El nombre TUBULINA, empleado para la
proteína principal de cilios y flagelos (véase más arriba), también se emplea
para designar la proteína de los microtubulos citoplasmáticos. La tubulina es
un dimero de 110 a 120.000 Dalton.
En los flagelos pueden identificarse dos
monómeros distintos, las tubulinas A y B. en la mayor parte de los casos, la
tubulina forma un heterodimero integrados por dos monómeros distintos pero de
peso molecular semejante (55.000 Dalton).
El
espacio de 8nm que se observa a lo largo del eje longitudinal de los
microtubulos mediante microscopia electrónica probablemente el apareciomiento
de dos monómeros de tubulina.
LOS
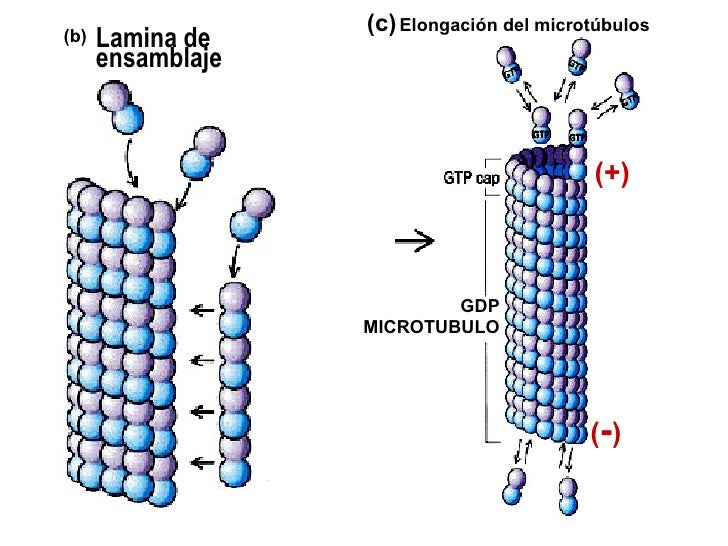
MICROTUBULOS SE ORGANIZAN A PARTIR DE DIMEROS DE TUBULINA, POLARIDAD.
El
montaje de los microtubulos a partir de los diámetros de tubulinas es un
proceso orientados y programado. Dentro de la célula existen sitios de
orientación, como centriolos, cuerpos basales de los cilios. A partir de los
cuales se dirigen la proliferación.
Estos
don los denominados centros organizadores de los microtubulos. Existen muchas
tubulinas polimerizada durante la interfase ( microtubulos citoplasmáticos) y
la metafase (microtubulos del huso), pero que disminuye en la profase y
anafase.
Dentro
de la célula los microtubulosestan en equilibrio con una fuente de moléculas de
tubulina libre. La polimeracion se favorece por la fosforillacion de la
tubulina por una proteiniquinasa dependiente del AMP cíclico. En las células
epiteliales cultivadas, el AMP clínico favorece a la formación de microtubulos
y la célula se vuelve alargadas. Hay pues una relación entre forma celular, número
y dirección de microtubulos y AMP cíclico.
La
agregación y disgregación de la tubulina es un fenómeno palarizado. En un microtubulo la agregación de los
dimeros de tubulina tiene lugar en un extremo mientras que el otro predomina la
disgregación. Si se trata una célula con colchicina se inhibe su agregación,
pero la disgregación continua, lo que lleva a la desorganización de
microtubulo. La agregación va a acompañada por la hidrolisis del GTP a GDP y la
falta de GDP detiene la agregación.
ROTEUNAS MICROTUBULARES ASOCIADAS. Mediante estudios de la
polomerizacion de tubulina se vio que los micrutubulos se organizan a 37º C y
se desorganizan a 5º C. si se purifican los microtubulos mediante varios ciclos
de montaje-desmontaje, sino que hay un 5% de otras proteínas, que genéricamente
se han denominado proteínas microbulares asociadas (MAPs). Se aislaron varias
de estas proteínas con procesos moleculares de 300.000 a 55.000.
LOS
MICROTUBULOS PUEDESN DETECTARSE POR ANTICUERPOS
El
aislamiento de tubulina permitio preparar anticuerpos específicos contra esta
proteína. Estos antecuerpos marcados por fluorescencia pueden usarse para
localizar microtubulos en el citoplasma de diversas células cultivadas. También
se prepararon anticuerpos contra proteínas de los mocrofilamentos, lo cual dio
base para que en el mismo tipo celular se estudiara la localización de los
microtubulos y los microfilamentos.
Los microtubulos citoplasmáticos, en una célula
cultivada, se disponen radicalmente a partir del núcleo y se dirigen en forma
recta o curva para terminar cerca de la superficie celular. Los microtubulos
desparecen por la colcemida ( un derivado de la colchicina) o por la acción del
frio, y reaparecen si la condición se revierte. Se observa que los microtubulos
se forman a partir de uno o más puntos focales cerca del núcleo, los que
corresponden a la regios centrosimica (centrosfera). Con la mitosis, los
microtubulos citoplasmáticos desaparecen y son remplazados por los que integran
el huso y los ásteres.
LOS
MICROTUBULOS CITOPLASMÁTICOS CUMPLEN DIFERENTES FUNCIONES
FUNCION
MECANICA. La
forma de algunas prolongaciones o
protuberancias celulares fue correlacionada con la orientación y distribución
de los microtubulos. Estos son considerados como un armazón o cito esqueleto
que interviene para modelar la forma de la célula y para distribuir su
contenido.
MORFOGENESISI.
El papel de
los microtubulos en la adquisición de la forma durante la direnciacion celular está
relacionado con su función mecánica. La formación de microtubulos desaparece al
completarse el alargamiento nuclear y es reemplazada por otros sistemas de
microtubulos que corren en forma axil.
POLARIZACION
Y MOTILIDAD CELULAR. La determinación de la palaridad intrínseca de ciertas
células también está relacionada con los microtubulos. En varias células
cultivadas se demostró que el tratamiento con colcemida determina cambios en la
motilidad.
CIRCULACION
Y TRASPORTE. Los
microtubulos puedes actuar así también como un sistema micro circulatorio para el trasporte de
macromoléculas en su interior.
TRASDUCCION
SENSORIAL. En
los receptores sensoriales es frecuente encontrar conjuntos de microtubulos, y
se ha considerado que intervienen de alguna manera en la transducción de las
diferentes formas de energía incidente.
RESUMEN
PROPIEDADES
DE LOS MICROTUBULOS
Los
microtubulos se observan en todas las células eucariontes, sean libres en el
citoplasma o bien como parte integrante de los cilios y flagelos.
Tienen
un diámetro externo de 25nm y varios micros de longitud y una pared de 6nm de
espesor que presenta 13 subunidades. La estabilidad de los distintos
micrutubulos es variable; los citoplasmáticos y los de huso sueles ser lábiles,
en tanto que los de los cilios y flagelos son más resistentes a los distintos
tratamientos. El componente principal es una proteína denominada tubulina,
heterodimero de 110.000 a 120.000 Dalton formando por dos monómeros son de
4x6nm y probablemente correspondan al enrejamiento de subunidades que se
observan en la parede tubular. La molecula de la tubulina presenta dos sitios
distintos por medios de los cuales se une a la colchicina y vinblastina,
respectivamente. El montaje de la tubulina para formar los microtubulos es un
proceso orientado y programado, en el cual los centriolos, cuerpos basales y
centromenos intervienen como centros de orientación.
En
la agregación interviene la hidrolisis del GTP a GDP y la polaridad de la
agregación puede ser demostrada, con el microscopio electrónico, mediante el
agregado de tubulina.
No hay comentarios.:
Publicar un comentario