Estructura,movimiento y origen de los cilios y flagelos
Estructura,movimiento y origen de los cilios y flagelos
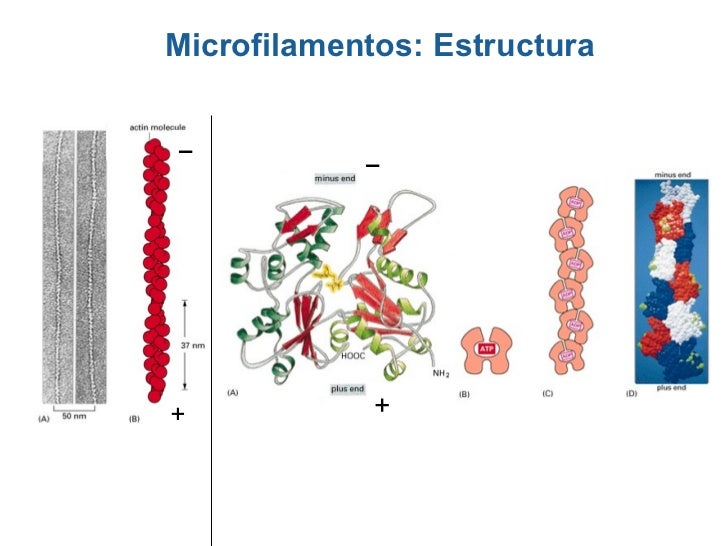
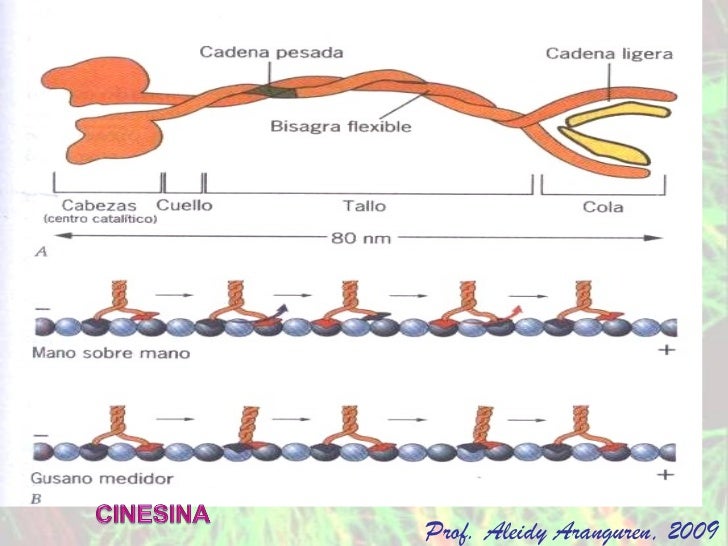
Son prolongaciones móviles que se encuentran en protozoarios poseen flagelos.los conocílios no deben ser confundidos con con lo estéreocilos,ya que estos últimos no poseen motilidad y tampoco tienen una estructura microtubular.El axonema es la estructura microtubular fundamental de los cilios,el cuerpo basal y las racillas ciliares.Distintas partículas morfológicas permiten distinguir permiten distinguir al subfibra A de la subfibra B de cada doblete.Los llamados brazos de dineina continen una AT pasa de alto peso molecular. Se piensa que la interacción entre tubilina y dienina esta base de mecanismos de contracción de cilio y flagelos.la nexina es otra proteina que une dobletes de microtubulos.El brazo externo Se asemeja a un gancho.mientras que el interno se conecta con la subfibra B adyacente ( este último constituye la conexión periférica).las conexiones radiales unen a cada subfibra A con la vaina central que contienen los microtubulos centrales.Los cuerpos basales o cinetosomas tienen la misma estructura que los centriolos. Un centriolo es un cilindro (0.2 x 0.5 m) abiertos en ambos extremos,aunque el extremo distal de los cuerpos basales se encuentra un placa ciliar que lo separa del cilio.la pared centroliar contiene tripletes de microtubulos, que desde él centro ala periferia son disgnados A,B y C. Los tú bulos A y B atraviesan la placa ciliar y se continúan con los correspondientes tubulos de cilio.